| Neural Stem Cells:
Implications for the Conventional Radiotherapy of Central Nervous System Malignancies. Barani IJROBP 2007;68:324 see anatomy picture
here
The
current understanding of neural stem cell (NSC) biology and
observations that neurogenesis endures in adult mammalian brain, provide a rich
interpretive framework for reexamining the results of past clinical studies in the
treatment of brain malignancies and suggest avenues for additional study. Spatially
discrete NSC niches may provide the means for improving therapeutic gain by enhancing the
differential effect on tumor and normal tissues of existing therapies. Radiotherapy (RT)
is ideally suited for the treatment of intracranial lesions, because it is not limited by
the blood–brain barrier and is able to conform to highly irregular target volumes.
For these reasons, RT is suitable for limiting the dose to the NSC
compartments, thereby preserving intrinsic brain repair capacity and limiting
treatment-related injury while delivering tumoricidal doses to sites grossly involved with
disease. The stem cell compartment contains a heterogeneous population of cells, some of
which are highly undifferentiated and endowed with a self-renewal capacity and an
extensive proliferative potential, that are responsible for the tissue homeostasis
throughout the life of an organism. NSCs produce an entire range of mature cell types that
are found in the brain (multipotency). This can be achieved by either asymmetric
divisions, by which a faithful copy of the mother cell, together with a more mature and
transiently dividing progenitor cell, is generated. Alternatively, the stem cell pool can
be maintained by an equivalent number of symmetric cell divisions, yielding either two
stem cells or two more mature progenitors. These descendants eventually give rise to
terminally differentiated elements of the mature brain. For example, transiently dividing
progenitors co-locate in neurogenic centers but retain multipotency that is restricted to
neuronal lineage cells. The resulting daughter cells then migrate throughout the brain
parenchyma and integrate as new interneurons in the cortical layers
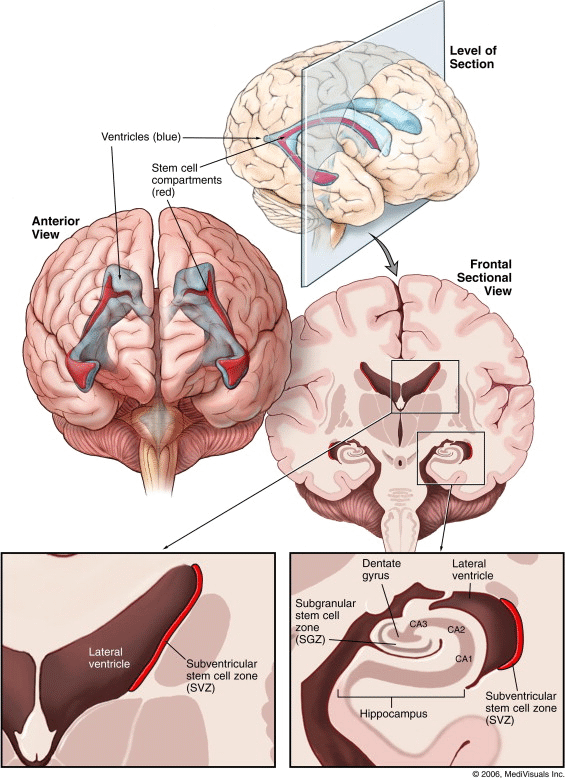
The largest neurogenic region in the adult mammalian brain is the subventricular
zone (SVZ), which is located between the lateral ventricle and the striatal
parenchyma (Fig. 1). A subset of SVZ cells
that express glial fibrillary acidic protein functionally serve as adult NSCs. These cells
have been shown to reconstitute the entire neurogenic structure when all other mitotically
active cells have been ablated. A similar hierarchical neurogenic
system is present in the subgranular zone (SGZ). The SGZ is located
between the hippocampal granular layer and the hilus and contains foci of proliferating
cells that are tightly associated with blood vessels. As in the SVZ, the SGZ
astrocytes that operate as NSCs give rise to transiently dividing precursors from which
new neuronal precursors are generated that migrate a short distance to functionally
integrate into the granule cell layer. The cytoarchitecture of these neurogenic niches is
complex, but it specifically contributes to the sustainability of the stem cell
compartment. The persistence of germinal regions and the presence of NSCs and transiently
dividing progenitor compartments in the adult CNS has important conceptual and practical
implications and reinforces the idea that essential components of
intrinsic brain repair capacity are confined to the SVZ and SGZ. The capacity of
NSCs to initiate and sustain repair, leading to preservation or reconstitution of
function, is highly dependent on the local microenvironment and the type of injury. It is
known, for example, that various inflammatory components can significantly impair
neurogenesis in vivo and limit full and long-lasting repair. This impairment, however, can
be reversed with the use of nonsteroidal anti-inflammatory drugs, such as indomethacin.
More importantly, elegant murine experiments demonstrated tropism of NSCs for areas of
brain pathology. NSCs have been shown to track down and destroy
migratory tumor cells and facilitate repair of tumor-related injury. For example,
animal studies using NSCs as delivery vehicles for tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL) and interleukin-12 resulted in collocation of NSCs and
tumor cells. Taken together, these data illustrate the uniqueness and importance of NSCs
and early progenitor cells in maintaining tissue homeostasis in the mature CNS. In murine models, a progressive decline in neurocognitive function
after irradiation was accompanied by an increase in hippocampal apoptosis, a decrease in
hippocampal proliferation, and overall decrease in neurogenesis, even at doses <2 Gy .
It is likely that postnatal neurogenesis mediated by the SGZ plays a critical role in
normal hippocampal function and that radiation damage leads to cognitive deficits. In
similar experiments, whole brain exposure to a single 10-Gy dose resulted in ablation of
hippocampal neurogenesis and preferential asymmetric differentiation along glial, not
neuronal, fates on transplantation of nonirradiated NSCs into the irradiated hippocampus.
These data have also demonstrated that the extent of inflammation correlated with the
radiation dose and, in turn, that this was associated with decreased neurogenesis. The
number of activated microglia, a proxy measure of inflammation, remained significantly
elevated up to 2 months after irradiation, suggesting a subacute or chronic time course.
The effects of radiation on neurogenic centers can be summarized
thus: (1) NSCs exhibit exquisite in situ radiosensitivity; (2) radiation can
directly depopulate NSC niches, causing immediate loss of NSC-mediated repair and
plasticity; (3) indirect effects of radiation are inflammatory in nature, dose dependent,
and capable of stunting neurogenesis even at low doses; and (4) the scope of NSC
dysfunction is age dependent, with greater effects noted in immature brains.
Implications for Therapy
Recent advances in basic neuroscience have shown NSCs to be the source of plasticity and
repair in the mature mammalian brain and have demonstrated that NSCs and CSCs share many
of the same phenotypic characteristics, even possibly the same parent cell. Given our
current inability to discriminate between these two cell types on the basis of their
biologic properties, treatments that are effective against CSCs are, therefore, likely to
also affect NSC viability and function. Thus, despite numerous clinical attempts, only
marginal therapeutic gains have been attained.
A therapeutic gain can be improved by either limiting treatment-related toxicity or by
improving tumor control; however, the combination of the two approaches is ideal. This
review has presented evidence to suggest that treatment-induced NSC dysfunction might be
responsible for the clinically observed toxicity. Although the selective preservation of
NSCs using biologic markers is clinically not yet possible, the
predilection of NSCs for the periventricular (SVZ) and hippocampal (SGZ) niches permits
spatial discrimination. Thus, NSC-preserving treatment aims to minimize, or altogether
avoid, dose delivery to both the SVZ and the SGZ, while appropriately targeting the tumor.
RT is ideally suited for this role. If NSC-preserving RT is delivered in the context of a
multimodality treatment that includes contemporary chemotherapy or targeted biologic
therapy, the benefits of NSC preservation could be ameliorated, given the inability of
these systemic modalities to selectively protect the NSCs.
The ability of RT to control a tumor depends on multiple factors, of which cellular
heterogeneity, repopulation, and distribution of disease are perhaps the most relevant to
treatment planning. As previously mentioned, the relative radiosensitivity or, conversely,
the radioresistance of tumor clones derived from a single parent cell can vary
dramatically. Therefore, a tumor’s response to irradiation cannot be simply
represented by a single tumor control curve, but by a range of dose–response curves
bounded by those that correspond to the least and most radiosensitive tumor clonogens.
Comparably extensive variation in cell kinetics parameters can also play a significant
role in the preservation and even expansion of tumor clones. Additionally, repopulation of
a previously treated tumor cavity with migrating CSCs can yield a clinical picture of
tumor recurrence or persistence, despite adequate initial treatment. It then becomes clear
that no single fraction size will effectively cover the range of radiosensitivities
represented by this variation and why hyper- or hypofractionation alone, or dose
escalation alone cannot effectively address tumor control without undue side effects.
Brain RT should, therefore, be delivered in a NSC-preserving manner with fractional doses
significantly >3 Gy and a total treatment time of <2 weeks—that is,
intensified, accelerated hypofractionation.
NSC preservation can initially be approached by defining the SVZ
as a strip of periventricular striatum generated by a 5-mm lateral expansion from the
lateral wall of the lateral ventricles as seen on T1-weighted magnetic resonance imaging.
Similarly, a 5-mm expansion of the hippocampal formation can safely delimit the SGZ.
Although the current dose limits are not firmly
known, the dose tolerance of the NSC compartments can be
estimated to be in the range of 10–20 Gy, with notable fraction-size dependence.
With daily fraction doses of 3, 2.5, or 2 Gy, the NSC compartment daily fraction doses
would likely need to be 100, 83, and 66 cGy, respectively. These guidelines provide a
useful starting point for additional investigations of the NSC and CSC paradigms, with
expected refinement of these estimates.
Conclusion
Dose-dependent preservation of NSCs, and thereby of the intrinsic brain repair mechanisms,
through judicious use of NSC-preserving RT has the potential to ameliorate
treatment-related toxicity. This could be particularly relevant in the pediatric
population and those with potential for long-term survival. Additionally, the combination
of NSC-preserving RT and differential fractionation according to disease burden has the
potential to further increase the therapeutic index. The treatment planning concepts
discussed in this review are amenable to implementation with current RT technology and are
worthy of further investigation. However, the modification of treatment portals outside of
the protocol setting is currently not warranted. Careful patient selection, definition of
failure patterns, and comprehensive evaluation of neurocognitive performance should be
central in future clinical investigative efforts. Studying the benefits in clinical
outcome of this approach, relative to conventional treatment planning, would be
appropriate for multicenter prospective evaluation.
|
{kind=link}